Research
 The Astrotactins
The AstrotactinsGlial-guided neuronal migration is a key step in the development of laminar architecture of cortical regions of the mammalian brain. The neuronal protein Astrotactin (ASTN1), a neuron-glial ligand during CNS glial-guided migration, was discovered in the Hatten Lab in 1988 (Edmondson et al, 1988). Recently we showed that a second family member, ASTN2, regulates the trafficking of ASTN1 during glial-guided neuronal migration (Wilson et al, 2010).
 Astn2 in Autism Spectrum Disorders
Astn2 in Autism Spectrum DisordersHuman genetic studies on ASTN2 reveal that mutations in ASTN2 are associated with developmental diseases, such as autism spectrum disorder (ASD) and schizophrenia. To study ASDs, we are focusing on human cells containing mutations in ASTN2. This project is using a novel genetic method, translating ribosomal affinity purification (TRAP), in mouse cells and in cells from human patients with a deletion or mutation in ASTN2. We are profiling gene expression changes in cerebellar granule cell progenitors or human IPSC cells differentiated into granule neurons and using bioassays developed in the Hatten Lab to test the effects of mutations in specific ASTN2 transcripts on neuronal migration and synaptic plasticity.
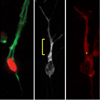 The Conserved Polarity Protein mPar6 Controls Glial-Guided Migration
The Conserved Polarity Protein mPar6 Controls Glial-Guided MigrationIn 2004, the Hatten Lab discovered that the conserved mPar6α polarity complex regulates neuronal migration by coordinating the movement of the neuronal centrosome and nucleus (Solecki et al, 2004). Recently we showed that mPar6 controls acto-myosin contractility by regulating the phosphorylation and activity of myosin light chain (MLC) and MLC phosphatase (MLCP) (Solecki et al, 2009). Our findings suggest that acto-myosin contractility in the proximal region of the leading process, rather than in a classical leading edge, generates the force needed for forward movement.
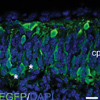 Lrp12/Mig13a, a Gene related to the C. elegans Migration Gene Mig-13 in Corticogenesis
Lrp12/Mig13a, a Gene related to the C. elegans Migration Gene Mig-13 in CorticogenesisDuring corticogenesis, the earliest generated neurons form the preplate, which evolves into the marginal zone and subplate. As the preplate separates, Lrp12/Mig13a positive neurons form a pseudo-columnar pattern, prior to moving to a deeper position within the emerging subplate layer. These changes in neuronal polarity do not occur in reeler mutant mice, revealing the earliest known defect in reeler cortical patterning (Schneider et al, 2010). We are currently generating conditional loss of function mutations to assay the role of this gene in both cortical and cerebellar development, and carrying out biochemical studies to define the binding partners of LRP12/MIG13a.
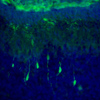 Gene Expression ProBiling of ES Cells using Translating Ribosomal Affinity PuriBication (TRAP)
Gene Expression ProBiling of ES Cells using Translating Ribosomal Affinity PuriBication (TRAP)To profile gene expression in cerebellar granule neurons at all stages of development and compare the biological signature of native granule cell progenitors with ES cells differentiated to a granule cell identity, we are currently using TRAP. We have established mES lines from Tg(NeuroD1-EGFP-L10a) transgenic mice that express the TRAP tag in cerebellar granule cells and differentiated them as we previously described (Salero and Hatten 2007). These experiments are revealing new developmental pathways critical for ES cell differentiation and providing a more rational basis for stem cell replacement therapy.