Multimedia

Vice President and Chief Scientific Officer, Howard Hughes Medical Institute
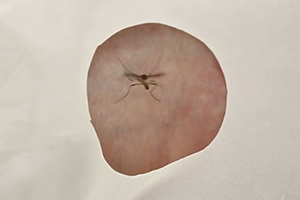
This animated video demonstrates current models of how the mammalian tongue recognizes and discriminates the different odor qualities: salty, bitter, sour, sweet, and umami (M.S.G.). A taste bud on the tongue is depicted. Different types of tastes activate defined taste cells that are tuned to a given taste quality. Most of the data leading to this model were generated by the group of Charles S. Zuker at UCSD.
Three-dimensional animation by Joe Alexander, Information Technology, The Rockefeller University, 2006.
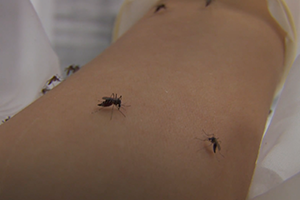
This animated video demonstrates current thinking about combinatorial coding in the nose. Odors are represented by different shaped objects floating in the air. Odors interact with olfactory sensory neurons that express different odorant receptors that are color-coded in the animation. Note that the blue odorant activates only three of the four different types of olfactory sensory neurons. Interestingly, different neurons respond differently to the same odor, showing strong, medium, and weak activation. Action potentials that result after odorant receptor activation propagate down the axon of each olfactory neuron and lead to differential activation of olfactory glomeruli in the olfactory bulb. Inhibitory interneurons will modulate this signal and transmit a filtered signal to higher brain centers.
Three-dimensional animation by Joe Alexander, Information Technology, The Rockefeller University, 2006.
An odor gradient of banana odor (isoamyl acetate) forms from the right side of the chamber. The chemotaxis behavior of single fruit fly larvae is tracked. These animals are manipulated to have very limited olfactory input. The animals colored green have only a single active olfactory neuron on one side of the animal. Those in blue have the same bilaterally symmetric olfactory neuron active on both sides of the animal, but all other olfactory neurons are silent. Note that the blue animals navigate more accurately in the odor gradient.
This data visualization by Jane Nisselson and Christine Yee is based on primary data from this article from the Vosshall Lab:
Matthieu Louis, Thomas Huber, Richard Benton, Thomas P Sakmar & Leslie B Vosshall. Bilateral olfactory sensory input enhances chemotaxis behavior. Nature Neuroscience 11:187-199 (2008).
Credit:
Data visualization by
Jane Nisselson, Virtual Beauty
Christine Yen, MIT Class of 2009, M.Eng 2010, Department of Computer Science
An odor gradient of banana odor (isoamyl acetate) forms at the far end of the odor chamber. The chemotaxis behavior of single fruit fly larvae is tracked. These animals are manipulated to have very limited olfactory input, such that one bilaterally symmetric olfactory neuron is active on both sides of the animal, but all other olfactory neurons are silent. The 2 spheres at the head of the larvae signal if the animal is heading left or right (depending on which one is red).
This data visualization by Jane Nisselson and Christine Yee is based on primary data from this article from the Vosshall Lab:
Matthieu Louis, Thomas Huber, Richard Benton, Thomas P Sakmar & Leslie B Vosshall. Bilateral olfactory sensory input enhances chemotaxis behavior. Nature Neuroscience 11:187-199 (2008).
Credit:
Data visualization by
Jane Nisselson, Virtual Beauty
Christine Yen, MIT Class of 2009, M.Eng 2010, Department of Computer Science
An odor gradient of banana odor (isoamyl acetate) forms at the far end of the odor chamber. The chemotaxis behavior of single fruit fly larvae is tracked. These animals are manipulated to have very limited olfactory input. The animals colored green have only a single active olfactory neuron on one side of the animal. Those in blue have the same bilaterally symmetric olfactory neuron active on both sides of the animal, but all other olfactory neurons are silent. Note that the blue animals navigate more accurately in the odor gradient. The 2 spheres at the head of the larvae signal if the animal is heading left or right (depending on which one is red).
This data visualization by Jane Nisselson and Christine Yee is based on primary data from this article from the Vosshall Lab:
Matthieu Louis, Thomas Huber, Richard Benton, Thomas P Sakmar & Leslie B Vosshall. Bilateral olfactory sensory input enhances chemotaxis behavior. Nature Neuroscience 11:187-199 (2008).
Credit:
Data visualization by
Jane Nisselson, Virtual Beauty
Christine Yen, MIT Class of 2009, M.Eng 2010, Department of Computer Science